レンスキーの研究グループは、細菌(ここでは大腸菌)を20年間世代交代させて、彼らの形質がどのように変化するかを調べました。
言うまでもなく無性生殖の細菌は、世代交代のスピードが速く、1日につき6~7世代繁殖すると言います。よってレンスキーの20年に及ぶ実験では、なんと45000回に及ぶ世代交代を観察できることになります。
この実験に細菌を使った理由は他にもあって、実験開始前のオリジナルの細菌や、任意の世代の細菌のサンプルを冷凍保存し、キープ出来ると言うこと。サンプルを再び解凍すれば、彼らは生命活動を再開してくれるので、実験前の細菌と、実験後の“進化”した細菌の形質を比較することもできますし、適応度を調べるために同一環境で暮らさせて競合させることもできます(この場合に備えて細菌にはあらかじめ遺伝子操作を施し、旧型と新型を色で区別できるようにしています、Ara+遺伝子は赤、Ara-遺伝子は白)。
レンスキーは、同一の遺伝子(クローン)の細菌を12のフラスコに分けて入れて、それぞれに独立した進化を促しました。
例えるならば、動物を遠く離れた12の大陸に別々に生息させたわけです。早くから他の大陸と独立したオーストラリアに住む動物に独特な固有種が多いのはこのためですが、それでも流木などで海を渡ってガラパゴス諸島にいっちゃう例もあるので、レンスキーの実験はそんな偶然すら許さない絶対的な隔離だと言えます。
レンスキーの実験は、個々のフラスコにグルコースと言う細菌の餌を入れて、細菌たちを育てるわけですが、増殖に伴い細菌の餌は次第に減っていき、食糧不足に陥ります。このような状態を停滞状態(プラトー)といい、大体グルコースは一日で尽きるそうです。
レンスキーのグループは一日経過しプラトーとなった細菌たちの一部(100分の1を無作為かつ正確に抽出して)を餌が入った新しいフラスコに移します。
この作業を毎日12の「フラスコ大陸」に別々に行い、12系列の細菌の進化を見届けるのです。つまりフラスコ大陸では1日ごとに、大豊作と大飢饉が繰り返され適度なプレッシャー、淘汰圧を与えている事になります。
20年後、7000回フラスコを取り換え=餌を供給し細菌が45000世代経過すると、程度の差はあれ、どの大陸でも打ち合わせでもしたかのように、同じ進化の傾向、細菌の大型化が見られたのです。
この実験で分かることは、進化のメカニズムはあくまでも「偶然」だとしても、生物は環境と絶えず「相互作用」をしており、変化は必然的に起こると言う事。
そしてその変化は、初期値やその環境が同じならば、それぞれのコロニーが辿った進化の細かな過程の差はともかく傾向も似通ったものになると言う事がこの実験から見て取れます。
・・・引用終わり。さて、この実験がなぜ小進化と大進化を結び付けられるのかというと、これはまごうことなき大進化の実験だからだ。
大腸菌をキリンに置き換えてみよう。キリンの平均寿命は野生下では15歳ほど。また大体4年ほどで成熟する。
ここでキリンは成熟してすぐ子どもを産んだとして、それが45000世代経過するには単純計算で4×45000=なんと18万年となる。
つまりレンスキーの実験はキリンを18万年間、世代交代させたことになる。もちろんこれは単純かつ強引なアナロジーに過ぎないが、大腸菌にとってはそれほどの長い年月が流れたということをイメージして欲しい。
そして大腸菌の各コロニーにおける平均の大きさの推移はグラフにすると程度の差はあれ大体このようになる。
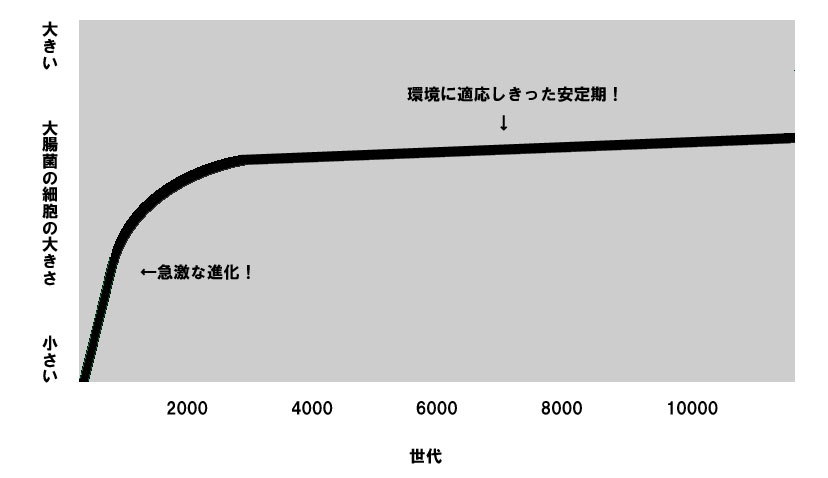
このグラフから解ること、それは細胞の大きさが大きくなる急激な進化は大体2000世代までで大体終了し、その後は緩やかになっていることだ。というかもうほとんど変化がない・・・
念のために言っておくが、これは1日分の食料が尽きたために起きたプラトーではない。10000世代という実験年数で言えば5年近い長期的な進化の傾向である。
つまりこの実験は、小進化のメカニズム(=自然選択)に則って毎日試行を繰り返した結果生じた大進化のグラフが、比例のような変化の割合が一定の直線的なグラフではなく、急激な変化とその後の停滞という断続平衡モデルを示す双曲線を描くことを立証したのだ。
そして大進化のグラフがすぐに安定状態に入ったことは何を意味するのか?これは「現段階ではこれ以上環境に適応する余地なし」ということかもしれない。
しかし、12のフラスコ大陸のコロニーのうち3番目のコロニーの大腸菌だけ、33100世代を境に急激に個体数密度を増したのである。まさに断続平衡モデルのように。
これは第3コロニーにおいて、今まで大腸菌が食べられなかった「クエン酸塩」を食べることができる大腸菌が突然変異で発生したからである。
しかしなぜ、どのコロニーの大腸菌も体の大きさは同じように大きくなっていったのに、クエン酸塩を食べるというブレイクスルーをやってのけたのは第3コロニーだけだったのか?
レンスキーによれば大腸菌がクエン酸塩を分解するために必要な突然変異が複数あったからだと言う。クエン酸塩を食べられるようになるためには、Aという突然変異とBという突然変異の両方が起きなければ食べられない。
つまり突然変異Aだけ起きても、突然変異Bだけ起きていてもクエン酸塩は全く食べられない。
ある時、第3コロニーの大腸菌は有益な効果は全くない突然変異Aを起こした(これはレンスキーのチームによって20000世代あたりで起きたことが確認されている)。この突然変異は中立的であってもなくてもいいようなものだったが、このことが突然変異Bを活かして受け入れる準備になっていた・・・
そして突然変異Bが起きた時、それは第3コロニーに新たな急激な変化をもたらしたのである。これこそまさに木村資生の中立進化!
つまりこの実験はなにがすごいかって、木村資生の分子レベルの偶然に作用される小さな中立進化が、ある時自然選択に大きく作用するような形質の出現をもたらし、そしてそのような小進化の繰り返しが、グールドの断続平衡的な双曲線のグラフを描いてしまうということがすべて分かるからだ。
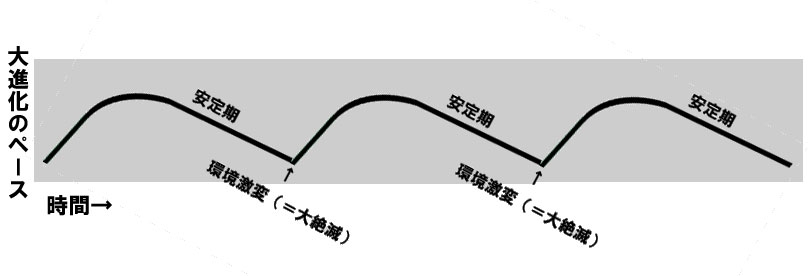
そして実際に地球で起きた38億年に及ぶ大進化はレンスキーの実験のように外的環境のルール(毎日グルコースを入れて群の100分の1を選別して・・・)が常に変わらないわけではないので、これよりもう少し複雑ではあるが、おそらくこのようになるのだと思う。
さてではそろそろ進化論についてのメカニズムをまとめて、長い旅を終えることにしよう。
①小進化のメカニズムが、絶えず小さな変化を繰り返しているのは事実だ。しかしそのほとんどは分子レベルの中立的な変異であり、自然選択にかけられるほどの顕在化はめったにしない。
②そして「大腸菌第3コロニーにおけるクエン酸塩の代謝」や「キリンのライバルの個体よりも少し長い首」など自然選択にかけられるほどの形質の変異は、その生物に適応か淘汰を促す。
③この形質が見事自然環境に適応した時、その生物集団がプールする遺伝子の頻度は移り変わり、生物種の平均的な形質(=外見)を微妙に変える。
④このプロセス(=漸進的な変化)を長い時間繰り返すことによって、生物の種は大きく形を変え、その環境にそれ以上適応の余地がないほど特殊化する(もちろんトレードオフの原理に基づいて)。
⑤こうなると生物の進化は緩やかになり安定期に入る。
⑥しかし定期的にやってくる隕石や地殻変動と言った環境の劇的な変化で、特殊化した生物は絶滅。彼らが埋めていた生態学的地位「ニッチ」に空きが出て、新たな急激な進化を1から促す土壌が出来る。これが大進化の断続平衡モデルである。
このまとめは進化の「時間軸」に基づいているが、実はまだまだ書き切れなかった進化のメカニズムはある。
例えばガラパゴスゾウガメやフィンチの種分化は、地理的隔離という「空間軸」に基づく進化だし、昆虫では地理的隔離によって生殖隔離(もう子どもが作れないほど形質がかけ離れ、ヒトとチンパンジーのような二つの種となってしまうこと)が起きる例も知られている。
またダーウィンも注目していた性淘汰(異性にもてるかどうかが進化に影響を及ぼすこと)や社会生物学の問題(働きアリなどの個体は利己的な生存競争をしてないぞ)をカットしてしまったのは、ほんとすいません・・・
このサイトで進化に少しでも興味を持ってくれたのなら、ぜひ『進化心理学入門』『進化学の挑戦』あたりをお勧めします。ではまた!