メンデルの遺伝の法則
言わずもがな。センター試験では遺伝は確か毎年出題されている。
最近の高校の教科書では、なんか優性の法則も教えていないらしい。いろいろ誤解を含む表現だからだそうな。その代わりにバイオテクノロジーのページがかなり増えた。
分離の法則
配偶子を形成する際、一対の遺伝子はそれぞれ分かれて別々の配偶子に入ること。
したがってRrの遺伝子を持つ親はRの配偶子とrの配偶子の2種類が作れる。
しかしRRの親はRだけを、rrの親はrだけをもつ配偶子しか作れない。そのため、この二つを交雑すると第二世代の遺伝子型はすべてRrになる。
優性の法則
対立遺伝子の働きには優劣関係があること。これは発現における優劣であって、形質における優劣ではないことに注意。
例えば、エンドウ豆の種子の形を丸にする遺伝子は、シワにする遺伝子に対して優先的に働く。
また完全に優性が発現せず中間雑種を作る場合がある(ピンク色のマルバアサガオなど)。これを不完全優性という。
独立の法則
独立の法則とは、それぞれの形質を決めている遺伝子どうしは、互いに干渉せず、それぞれ独立して配偶子に入るという法則。
例えばエンドウ豆で、種子の形を丸かシワにする遺伝子Aa、種子の色を黄色か緑にする遺伝子Bbは、別々に配偶子に入るため、AaBbという遺伝子を持つ親が作る配偶子は、AB、Ab、aB、abの4種類が等しくできることになる。
よって、種子がシワな場合は必ず種子の色は緑に決まっている・・・みたいなことがない。
しかし、染色体が連鎖をしている場合は、この法則は成り立たない。
連鎖とは遺伝子が同一の染色体上にある状態をいい、そのため連鎖している遺伝子は染色体が切れない限り行動を共にするため、独立の法則が成り立たないというわけだ。
例えば、エンドウPにおける1対の相同染色体の片方にAとB、もう片方にaとbが連鎖している場合、Pが作る配偶子はABとabの2種類(AB:ab=1:1)だけなので、エンドウP同士を交雑してできる第二世代のエンドウF1の表現型はAB:ab=3:1となる(大文字を優性だと仮定した場合)。
複対立遺伝子
ヒトのABO式血液型は、9番染色体に座位する3つ以上の対立遺伝子によって調節されている。
夫婦の血液型がそれぞれA型とB型で、その遺伝子型が判明していない場合は、夫婦の遺伝子型のパターンはAA×BB、AO×BB、AA×BO、AO×BOの4通りの可能性があり(A型はAA、AO、B型はBB、BOのそれぞれ二通り考えられるため)、生まれる子どもの血液型の確率はそれぞれ以下の通りである。
AA×BB
AB型の子ども:100%
AO×BB
AB型の子ども:50%
B(BO)型の子ども:50%
AA×BO
AB型の子ども:50%
A(AO)型の子ども:50%
AO×BO
AB型の子ども
A(AO)型の子ども:25%
B(BO)型の子ども:25%
O(OO)型の子ども:25%
この4つのパターンの子どもが生まれる割合を全て足すと、AB型56%、A型18.7%、B型18.7%、O型6.2%となる。
DNAの構造
DNAは正式名称をデオキシリボ核酸(デオキシリボヌクレイックアシッド)という。
その特徴は以下の3つである。
①DNAのほとんどは核の中に含まれる(例外:ミトコンドリアのDNA)。
②ひとつの体細胞の核に含まれるDNA量は生物種によって一定であり、体細胞の種類によって異なることはない。
③減数分裂によってできる生殖細胞ではDNAの量は体細胞の半量になるが、受精によってもとの量に戻る。
DNA分子は、ヌクレオチドという単純な単位の反復である。
このヌクレオチドの並びを塩基配列といい、この順番によって遺伝情報がコードされている。つまりDNA分子は遺伝子の正体であり、これは1952年のハーシーとチェイスのバクテリオファージの実験によって立証された。
しかしDNA分子すべてが遺伝子であるわけではなく、RNAという分子に転写される部分(全体の5%ほど)が遺伝子であるとされている。
DNAの構造は、リン酸を介してヌクレオチドが複数つながった1組のポリヌクレオチド鎖が反平行に対合し、らせん階段のようにねじれている(二重らせん構造)。
この構造は1953年にワトソンとクリックが明らかにした。
らせん階段のステップは、1ひねり分のらせん(1ピッチ)で10段あり、そのステップを形成する塩基には、アデニン、チミン、グアニン、シトシンの4種類がある。
この4種類の塩基は、それぞれ結合する相手が決まっており、Aは必ずTに、Gは必ずCに結合する。こうして、階段の1つのステップは、2つの塩基が水素結合を起こして作られる。
この塩基対ルール(シャルガフの規則)によって、DNAは一本のポリヌクレオチド鎖さえあれば、相補的なもう一本のポリヌクレオチド鎖の配列を割り出すことができる。
つまり、二重らせんをほどけば、片方の鎖を鋳型として新しい鎖を作ることができるというわけである。
DNAがコードしている遺伝情報はRNAという分子にコピーされ(転写)、このコピーを元に情報が読み取られ、タンパク質が合成されていく(翻訳)。
このDNA→RNA→タンパク質という一方的な流れをセントラルドグマというが、現在ではRNAからDNAが作られたり(逆転写)、わりとインタラクティブ性があったのでこの中央教義は崩壊の憂き目に遭っている。
突然変異の影響
一つの塩基が置き換わるタイプの突然変異を点突然変異といい、以下の3種類がある。
ミスセンス突然変異は、1つの塩基が変わったことで、異なるアミノ酸が運ばれてしまい、それゆえにタンパク質の構造が変わってしまうような点突然変異である。鎌形赤血球貧血症の原因がこのミスセンス変異である。
ナンセンス突然変異は、終止突然変異ともいい、アミノ酸をつないでタンパク質を作るのをやめてしまうような、点突然変異である。したがって非常に影響が大きい。
サイレント突然変異は、非表現突然変異ともいい、タンパク質におけるアミノ酸の配列に影響を及ぼさない点突然変異である。なぜこのようなことが起こるのかというと、1種類のアミノ酸をコードするコドン(ひとつのアミノ酸をコードする3つ続いた塩基配列のこと)が複数あるからである。例えば、UUUでもUUCでもフェニルアラニンというアミノ酸をコードするので3番目のUがCに変異しても作られるアミノ酸は変わらない。
一つの塩基が失われたり、挿入されると、変異箇所以降の塩基配列が全てずれ、大規模な読み込み枠の移動であるフレームシフト突然変異が起こる。
この影響は点突然変異よりも大きく、終わるべきところでアミノ酸の読み取りが終わらなくなったり(ポリAテールというmRNAの末端まで読み取ってしまう)、本来よりも早い場所でアミノ酸の読み取りが終わってしまい、タンパク質の長さが短くなってしまったりする。
しかしこれが3つの塩基が失われたり、挿入されると、その影響は比較的小さくなる(塩基3つでひとつのアミノ酸をコードするため)。
またサプレッサー突然変異は一回目の突然変異の効果が二回目の突然変異によって打ち消されてしまうような突然変異を言う。
タンパク質の構造
タンパク質はアミノ酸の列が複雑に折りたたまれてできている。
アミノ酸は全20種類で種類はそこまで多くはないものの、つながる順序や個数は決まっていないため、タンパク質の種類は100億種類以上ある。
以下の図はタンパク質を構成するアミノ酸の構造である。
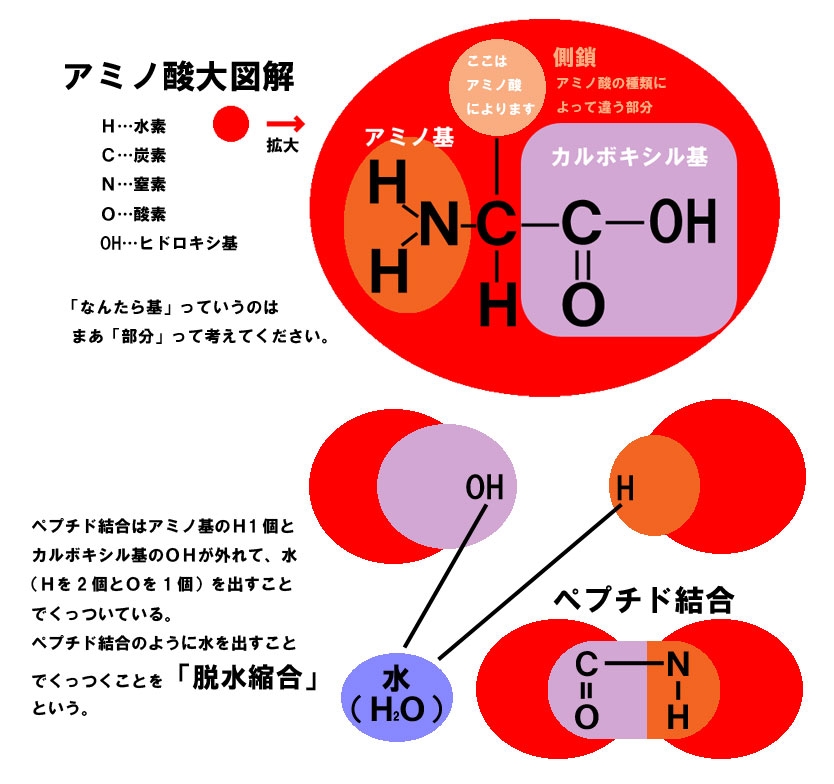
タンパク質の構造は以下の4つのレベルが存在する。
一次構造
何百個ものアミノ酸がペプチド結合でつながり、ポリペプチド鎖を作る。
ポリペプチド鎖の中には、お互いを引っ張ったり(引力)、反発したり(斥力)するアミノ酸が含まれているため、この配列がタンパク質のより高次の構造や、機能を決定する。
二次構造
ポリペプチド鎖が折り畳まれたもの。
代表的な折り畳み方には、らせん状のαヘリックスと、2本のポリペプチド鎖が平行で結合しジグザグ状に折れているβシートの二種類がある。
二次構造はCO基とNH基の水素結合で維持される。水素結合は単独では弱いが、スイミーのようにたくさん集まると非常に強い力になる。
αヘリックスもβシートもR基が外に突き出ており、この部分がタンパク質の科学的な性質を決めている。
ちなみに毛や爪などのケラチンは全てαヘリックス、絹の繊維を作るフィブロインは全てβシートで出来ている。
三次構造
すべてのタンパク質は、二次構造がさらに複雑な形に折り畳まれて出来ている。これを三次構造といい、二次構造の様々な場所が互いに引きつけ合うことで折り畳まれる。
最も強い結合は、隣接するシステインがジスルフィド結合でつながったもので、他にも、疎水性の相互作用、弱いイオン結合、水素結合などがある。
ちなみにパーマは髪の毛(ケラチン)のジスルフィド結合の位置を変えてかけるので、一度かけるとなかなかなおらない。
四次構造
酵素のようなタンパク質は三次構造で完成するが、多くのタンパク質はポリペプチド鎖がさらに集まって出来ている。
例えばヘモグロビン分子(C3032H4816O872N780S8Fe4)は、4つのポリペプチド鎖がサブユニットとしてくっついてできた球状のタンパク質である。
各サブユニットの中心にはヘム鉄があり、これが酸素とくっつくことで、赤血球は酸素を運搬している。
このような非タンパク質性の部分を補欠分子族といい、これを持つタンパク質を複合タンパク質という。
タンパク質の変性
タンパク質は複雑に折り畳まれることによって活性のあるR基の配置が決まり、それに伴って、そのタンパク質特有の化学的性質が決まる。
タンパク質の変性とは、この構造が消失してしまうことを指す。これによりタンパク質の機能も消滅し(失活)、一度変性したタンパク質はほとんどの場合、元には戻らない。その理由は、たとえアミノ酸配列はそのまま残っていたとしても、二次構造、三次構造を作る結合が変化してしまうからである。
タンパク質の変性を起こす原因は以下のものがある。
①強酸・強アルカリ
イオン結合を切断して、タンパク質を凝固させてしまう。
さらに強酸・強アルカリに長くさらすと、一次構造(アミノ酸のペプチド結合)も崩壊する。
②重金属
R基のカルボキシ基と強く結合することでイオン結合を切断、タンパク質の電荷を減らす。
③熱および放射線
原子に加わるエネルギーを単純に増やし、タンパク質の様々な結合を切断する。
④界面活性剤および溶剤
非電極基(電荷を持たない官能基)と結合し、水素結合を切断する。
ちなみに官能基とは有機化合物の性質を決める部分のことで、別にエロス的なものじゃない。