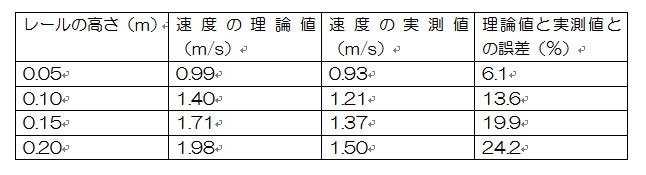
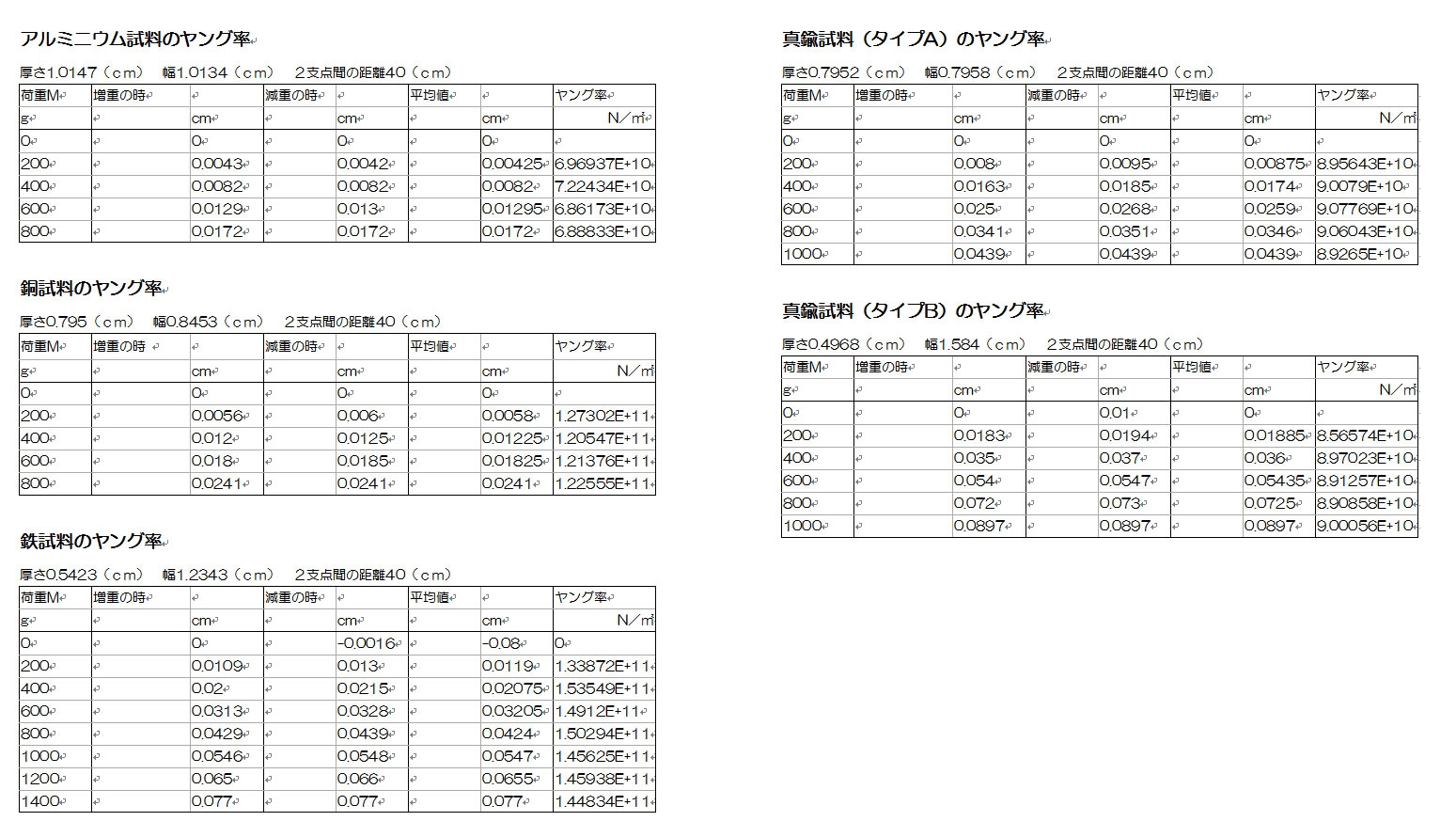
中学校で習う電磁誘導の法則(エルステッドの法則によって磁石が電流から力を受けるならば、逆に電流も磁石から作用反作用の法則によって力を受けるはずだというやつ)を実験で確認するだけのボーナス課題だが、中学校ではなかなか教えてもらえないフレミングの右手の法則や、検流モーター、相互誘導とインダクタンスなどが追加されている。
検流計の仕組み
メーター類もモーター同様、電磁誘導の法則によって針が振れるようになっている。メーター類に電流が流れると、位置を固定された磁石に挟まれたコイル内部に電流が流れることで磁界が発生し、この時できるコイルの磁界と磁石の磁界との反発力、もしくは引き合う力によって、コイルが動き出し、そのコイルに取り付けられた針が振れる。
針の振れる大きさは電流の大きさに比例するために、針の振れ幅によって電流の大きさを調べることができる。
フレミングの左手の法則と右手の法則
フレミングの左手の法則も右手の法則も、左手の親指と人差し指と中指を使って、電磁誘導現象における、電流(中指)、磁界(人差し指)、力(親指)の向きを表すものである。
ただフレミングの左手の法則は電流と磁界の向きから定まる電磁力(力)の方向を、右手の法則はコイルを動かす(力の)方向と磁界の向きから定まる、誘導電流の方向を示している。
ファラデーの法則とレンツの法則
電磁誘導現象によって発生する電流を誘導電流という。ファラデーの法則によれば、この誘導電流が流れる強さ(e)は、コイルを通る磁束の時間あたりの変化量(コイルと磁石を近づけたり遠ざけたりする際の速さ)、と磁束の強さ、そしてコイルの巻数および磁界の存在する媒質に比例する。
ファラデーの法則が誘導電流の強さについての法則であるのに対し、レンツの法則は誘導電流の向きについての法則である。
レンツの法則によれば、誘導電流の流れる方向は、コイルを通る磁束の変化を妨げる向きに誘起される。
また、ファラデーの法則とレンツの法則をまとめて電磁誘導の法則という。
検流モーター
直流モーターでは、固定された磁極に挟まれたコイルに流れる電流の向きを整流子によって半回転ごとに切り替えることでコイルを決まった向きに回転させていたが、検流モーターでは逆にコイルの位置を固定し、コイルの中の磁石を回転させている(インバーターモーターっぽい)。
検流モーターのコイル位置は固定されているため、電流の向きは端子をつなぎ変えない限り変化しない。
つまり、検流モーターと直流モーターでは、原理が同じでも、時間的に切り替わる要素が検流モーターでは磁石の向き、直流モーターではコイル(に流れる電流)の向きと異なっており、電流に対して磁界を変えることと、磁界に対して電流を変えることは、どっちも同じ結果であるということが実験を通して具体的に確認することができる。
次の図は検流モーターを上から見たものである。
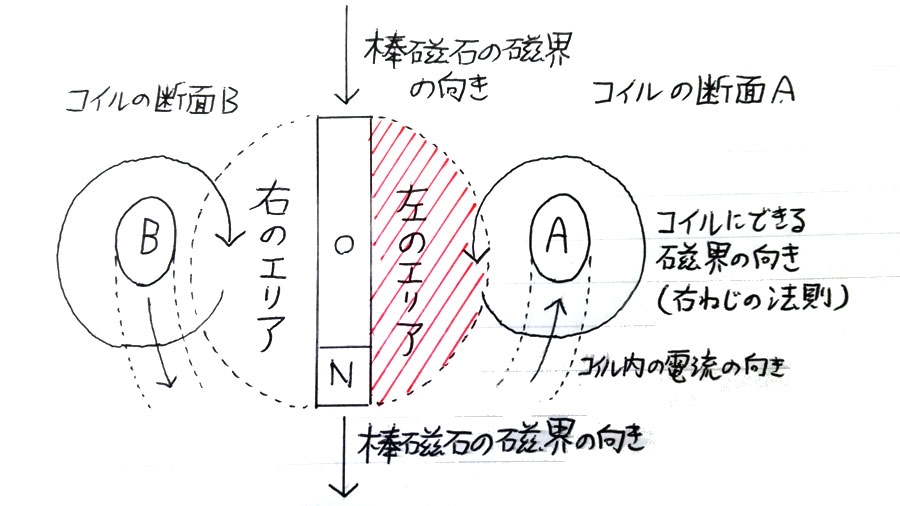
左右のコイルに流れる電流によって発生する磁界の向きは、右ねじの法則からAの箇所では反時計回り、Bの箇所では時計回りであるため、コイルに電流を流し続けると、磁石部の向きはN極が下、S極が上で静止する。
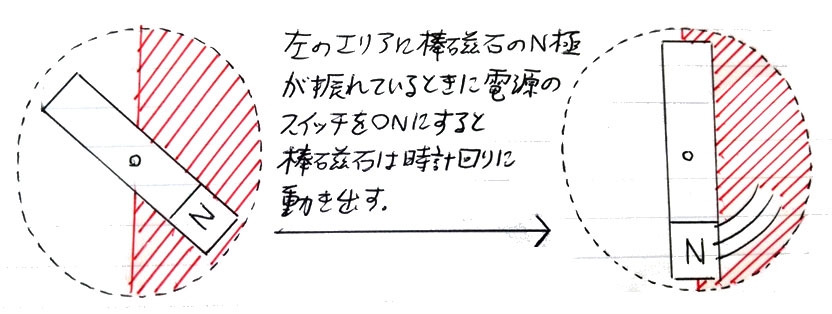
この状態からコイルに流れる電流をスイッチを連打することで点滅させると、磁石部が小さく振れ出すが、その上でさらに、タイミングよく磁石部のN極が図の右エリアに振れた時にスイッチをオン、図の左エリアに振れた時にスイッチをオフにすると、磁石部は時計回りに回転をはじめ(※)、逆に磁石部のN極が図の右エリアに振れた時にスイッチをオフ、図の左エリアに振れた時にスイッチをオンにすると、磁石部は反時計回りに回転をはじめる。磁石部の回転の勢いが強くなり、慣性で半回転するようになると、スイッチを点滅させるタイミングを間違わない限り、磁石部は一定の方向で回転を続けるようになる。
※検流モーターにおける磁石部の回転
相互誘導
電気的に接続されていない二つのコイルが磁束の変化によって相互作用をすること。
これにより片方のコイルに電気を流すことで、もう一方のコイルに誘導電流を生み出すことができる。また、その誘導電流の大きさはコイルの巻数で調整が効くため、電柱なんかのトランスはこれを利用している。
①検流モーター、手持ちコイル、電池ボックスを付属のケーブルで接続する。
②鉄心を手持ちコイルに通す。
③検流モーターの磁石部が、コイルと並行になるように配置する。
④電池ボックスのスイッチを押したままにする。すると、電池ボックスに接続されている手持ちコイルに電流が流れることになる。これにより、検流モーターに接続されている手持ちコイルに誘導起電力が発生し、その磁界によって磁石部が力を受け動き出す。
⑤わずかに動いた磁石部は、地磁気によって元に戻る。
⑥磁石部が元の状態(コイルに並行)になったときに電池ボックスのスイッチを離す。すると磁石の動きが加速する。
⑦④~⑥の操作を磁石部が半回転するまで続ける。
⑧磁石部が半回転したときに、スイッチを次の状態(押したり離したり交互にする)にして、磁石部が回転するようにスイッチを操作する。
コイルの中に鉄芯を入れると鉄芯が電磁石になるため、一次コイルの磁束の変化が、二次コイルに伝わり、二次コイルからの誘導電流によって検流モーターが動くが、鉄芯を入れないと一次コイルの磁束の変化が二次コイルに伝わらず、誘導電流が発生しないため検流モーターは動かない。
ペットボトルロケットを用いて放物線運動を考える
この班だけ神々の遊びがやれないのは可哀想、という優しきアシスタントの先生(教授の先輩)のお情けで、なんと急遽、振り子の重力加速度測定から予定変更。
こちら管制センター、フライトチェック完了、GOかNOGOか判断せよ、点火シークエンスを開始する、カウントダウン、これより打ち上げを実行・・・!(※たのしい)
ちなみに流体がない宇宙空間では別に翼はいらないのでつけない。じゃあどうやって姿勢を制御するんだ、と先生がJAXAに直撃したところ、コンピュータでスラスターのノズルの向きを調整しているのだという。
実験概要
ペットボトル型ロケットを用いて、放物線運動を再現し、その軌道を表す方程式で軌道計算を行い、軌道計算に用いたロケットの発射角度ならびに初速などの条件で打ち出したとき、実際と計算によるシュミレーションとを比較検討する。
このとき、ペットボトルが描く放物線運動に影響をあたえる因子について解明する。一方、その因子が一つではないことは容易に想像できるが、それだけに注意深く実験することが要求される。
実験器具
ペットボトル型ロケット
ロケットランチャー
空気入れ
スピードガン
レーザー放射距離計
給水用ペットボトル
計量カップ
角度定規
実験方法
①ロケットの後ろの栓をゆるめ逆さまにして計量カップで150ccの水を入れる。(BGM:キューピー三分クッキング)
②ペットボトルロケットをロケットランチャーにセットする。(注意:ランチャーの蝶ネジをゆるめ傾斜台をロケット発射角度に調整し、蝶ネジを締める。この時、ロケットの水がこぼれないようにランチャーを傾けてロケットノズルをジョインターにカチッと音がするまで確実に差し込む)
③ロケット内部に空気入れを使って空気を入れる。自転車用空気入れをジョインター・エアバルブにセットする。この時空気漏れがないことを確認。空気入れは15回ポンピングする。
④ロケットの発射はリモコンワイヤーを使って発射する。(注意:発射の前にロケットの前方・後方に人や動物がいないことを確認する。発射するときはランチャーの横方向に離れる。後ろにいると水がかかる場合がある。
⑤発射角度は45°、60°、70°の3つの角度について実験する。その時、角度は分度器で出来る限り正確に角度を決めセットする。ロケットの初速度を斜め前方からスピードガンを用いて測定する。
実験結果
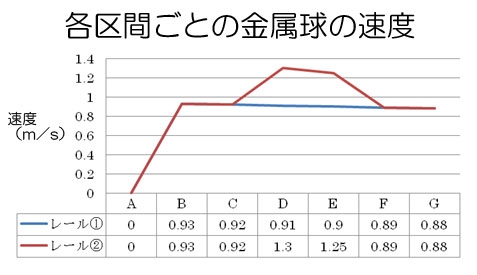
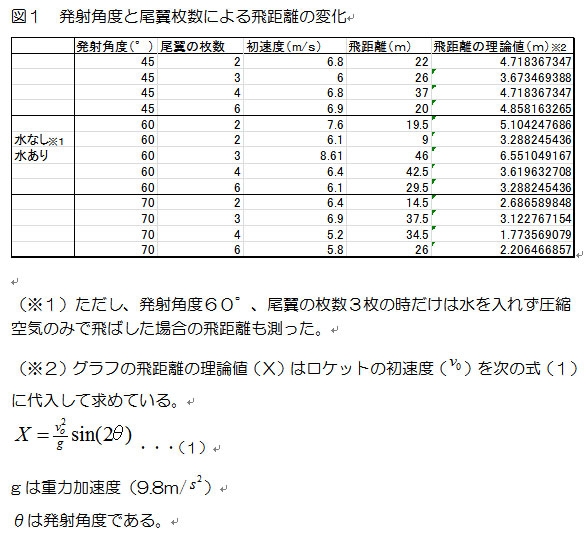
ペットボトルロケットに詰め込む水の量と空気圧を一定にした時のロケットランチャーの飛距離を、発射角度とロケットの尾翼の数ごとに以下のグラフにまとめた。
考察
本実験は放物線運動についてペットボトルロケットを用いて理解することを目的としている。
ロケットの到達距離に影響を与える要因は、ペットボトルに入れる水の量と圧縮空気との関係で決まるロケットの初速度、ランチャーの発射角度、尾翼の数といったロケットの形状、さらに打ち上げ場所と方向によって決まる風の影響などが考えられる。
以下、それぞれの要因について考察する。
①圧縮空気と水の割合
まず、ペットボトルロケットを飛ばすために圧縮空気と水を入れる理由について考える。ペットボトルロケットに水を入れ、空気入れでさらに圧縮空気を入れるとペットボトルの内部の圧力が高くなり、この状態で栓を抜くと高圧の空気が水を押し付けることで、水が栓から勢いよく噴射し、作用反作用の法則によってペットボトルロケットが打ちあがる。
すなわち水はロケットの推進剤の役目を果たしているのだが、この水の量が多すぎるとペットボトル内に入る圧縮空気の量が減り、逆に水の量が少なすぎても、推進力が低下しやはり飛距離が落ちてしまう。
実験では発射角度が60°、尾翼の枚数が3枚での打ち上げ実験の時に、水と圧縮空気を入れた場合と、圧縮空気のみの場合で飛距離を比較した。(グラフの※1)その結果、初速度には大きな変化はなかったものの、飛距離は5倍以上の差があった。
この原因は、圧縮空気のみの場合は瞬間的な推力しか生まれず、初速度のみで飛距離が決まってしまうが(注1)、水を入れた場合は水の粘度が空気に比べて高いために、ロケットが発射した(栓を抜いた)後も水が噴射されロケットが加速するからである。
また、ロケットが飛んでいく推力はロケットが噴射する物質の速度と質量に比例するため、質量が大きい水を入れたほうがロケットの飛距離はあがるが、あまりに重い燃料を積むとロケット自体の重量が大きくなり、飛距離は落ちるので、ロケットに積み込む推進剤の適切な重量が存在する。実際、宇宙ロケットの打ち上げでは、空になった燃料タンクを飛行途中で切り離すことで、ロケットの重量を軽くさせ推進力を維持している。
よって発射前の質量と、燃料を使い切った発射後の質量の差が大きいほどロケットは遠くに飛ぶということになる。これは作用反作用の法則で、軽いものと重いものが互いに押し合った場合、軽いもののほうが遠くに飛んでいくことをイメージすると理解ができる。
注1:式(1)がそれで、物体を飛ばす際の角度と、初速度のみを考慮している。このモデルは、野球でピッチャーがボールを投げるといった、瞬間的な推力で飛んでいく物体の飛距離を求める際に使われる。(ピッチャーの手を離れたあとのボールは推進力を持たないため)
②尾翼の数
大気圏では空気や風の影響を受けるため、飛行機やロケットの機体の方向や姿勢を制御する尾翼の役割は非常に大きい。
本実験ではロケットの飛距離に尾翼がどれだけの影響を与えているかを調べるために、尾翼の数が異なるロケットを同じ条件で発射してそれぞれの飛距離を比較した。
図1の結果では、尾翼の数が2枚、すなわち垂直尾翼がない状態だと、機体の姿勢が安定せず、きりもみ状態となり飛距離はでなかったが、これに垂直尾翼をつけた、尾翼の数が3枚のロケットでは(45°の場合を除き)最も飛距離を出すことができた。さらに尾翼の数を増やしたロケットも打ち上げたが、尾翼の数が3枚の場合と、あまり飛距離の差は見られなかった。特に尾翼が3枚と4枚の飛距離はほぼ同じであった。尾翼の数が6枚の場合は、尾翼の数が3枚と4枚のロケットよりも若干飛距離が落ちたが、これは尾翼の数を増やしたことでロケットの重量が増加したことが原因だと思われる。
したがって、ロケットの尾翼の数は2枚よりも3枚の方が飛距離は伸びるが、3枚以上になると飛距離にあまり影響を与えず、なおかつロケットの重量を考慮すれば3枚が最も効率的であると結論付けられる。
③その他の要因
まず、圧縮空気を入れた際の空気漏れが考えられる。実験では空気入れに取り付けたジョインター・エアバルブがゆるかった。
次に、風の影響が考えられる。実験時にはかなり風が吹いていたため、たとえば、発射角度45°における尾翼の数が3枚のロケットと4枚のロケットの飛行距離の結果だけ、ほかの角度の結果と異なり、尾翼の数が4枚のロケットのが3枚のロケットの飛行距離を大きく上回っているが、これは4枚のロケットを打ち上げた際に、東向きの強い追い風が吹いていたことによるものと考えられる。また、発射角度が70°の実験の際には北向きの風が吹き、発射されたロケットの方向が大きく北によれていた。
三番目は、ランチャーの発射角度である。理論上では45°の時にもっとも飛距離が伸びるはずなのだが、実験では発射角度が60°の時に最も飛距離が伸びた。これはランチャーの傾斜台の角度が積んだロケットの重みで変化し、設定以下の角度になってしまったと考えられる。つまり発射角度45°の場合は45°以下、60°の場合は60°以下で発射された可能性がある。また、角度45°で最も飛距離が伸びるモデル(式1)では、発射する高さや、打ち上げる物体の形状、温度や気圧といった気象条件を考慮していない。したがって、打ち上げ角度45°よりも60°の方が、打ち上げたロケットの高度は大きいため、上空の気圧の関係(注2)で理論値よりも飛距離が伸びた可能性も考えられる。
最後に、ロケット自体の形状のゆがみが考えられる。ペットボトルでできているために機体が変形しやすく、またロケット後部や尾翼の取り付け部分が痛み、向きが変わりやすくなっていた。実験中に補修をしたが、これも飛行距離に少なからず影響を与えたと思われる。
注2:高度が上がると気圧は下がる。飛行距離に影響を与える空気抵抗は空気密度に比例するため、気圧や湿度が低いと飛行距離は伸びると考えられる。