あの死闘から2日後・・・(早い)
二度とスロットには手を出すまいと誓った私だったが、店側がまた新たな沼要素を繰り出してきました。

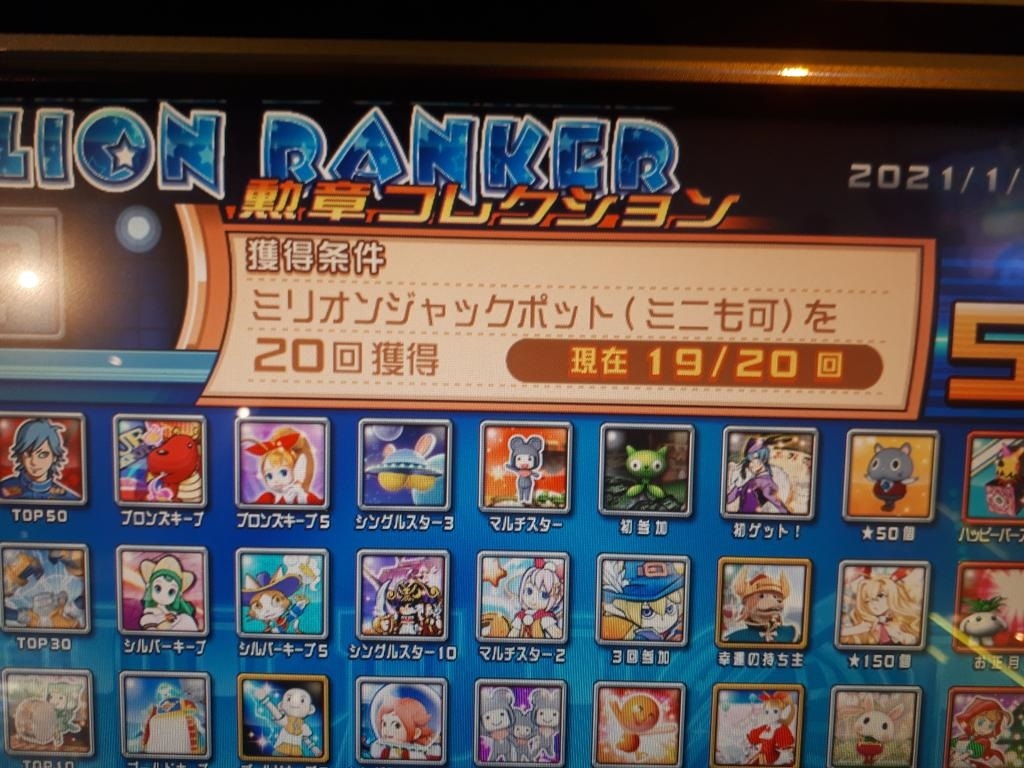
その名は、プレミアムチャンス!!その存在こそ噂には聞いていたが、2年以上このゲーセンに通っていて初めて出現した、3分間だけセブンラッシュに行く確率が上がるという大チャンスモードである!
今まで一度も見たことがなかったということは、店側がこのチャンスを出現させる設定をしていなかったんだろうけど、フィーチャープレミアムで大負けしたこのタイミングで設定を変えるとは、皮肉なもんだぜ。もしかして、この前のインカムのすごさを見て、向こうも味をしめたか??
しかし、私もメダルを1/3も失ってしまった以上、セブンラッシュの確率がアップするとは言え・・・やるけど。



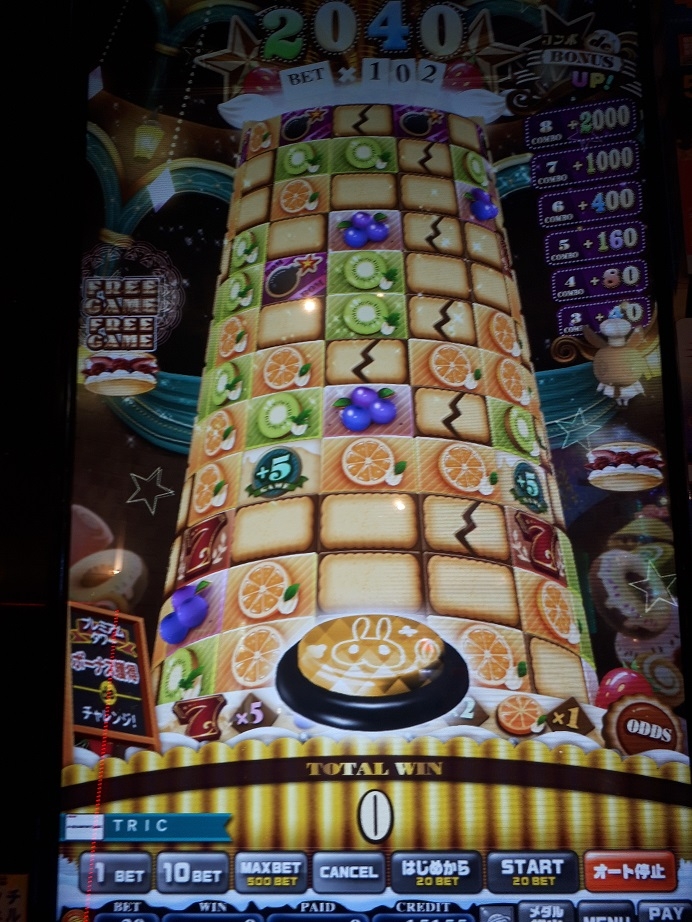
セブンラッシュに一度も行かねー!!!マックス500枚ベットで回しちゃったじゃねーか!
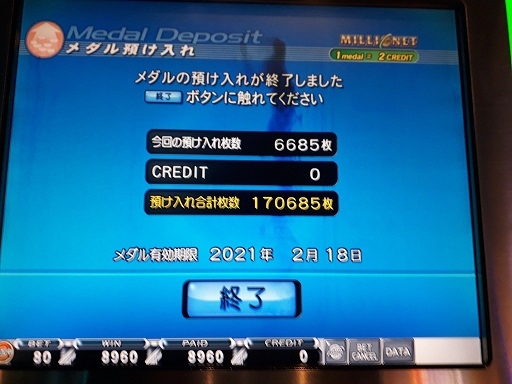
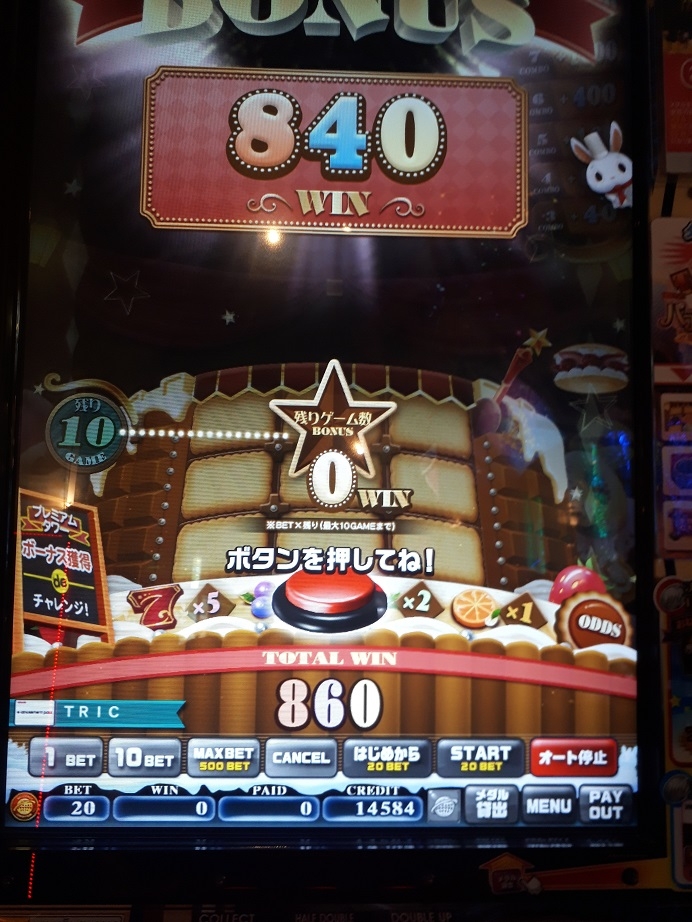
たった3分で10000枚減りました。なんつー詐欺モードだ・・・すると、プレミアムチャンスが終わってすぐミリオンジャックポットが始まり・・・

神にふさわしい枚数が戻ってきました。でもこれ、たまたまミリオンジャックポットがここまで溜まっていたからいいものを、1500枚くらいだったら爆死確実だったよな・・・
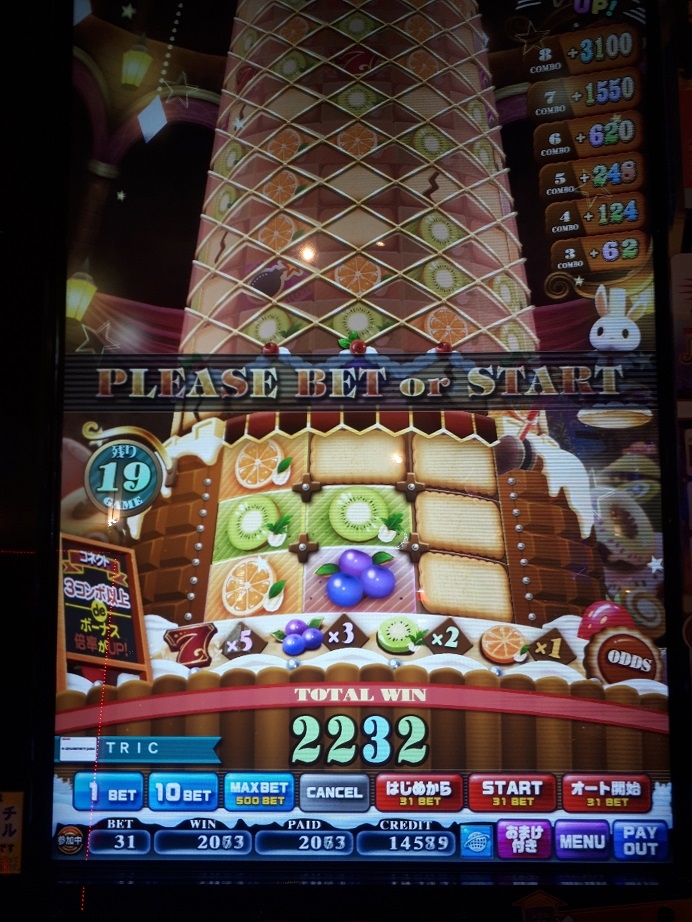
その後もプレミアムチャンスは度々発生するようになり、今度はちょっと疑ってかかって100枚ベットで回すものの・・・



やはり、セブンラッシュは出ず。なんだこれ。でも、まあ10倍配当は少なくとも出してくれるので、メダルを増やすチャンスではあります。
ちょっと悔しいので、今後もフィーチャープレミアム周辺で様子を見ながら、プレミアムチャンスで500枚ベットのセブンラッシュ(夢)を狙ってみたいと思います!
なんか、この前は、夕方の6時と8時前後に出てきたから、時間帯が決まっているのかも。だとしたら狙いやすいんだけど。始まってから慌ててログインすると、暗証番号入力やメダルを転送もろもろで1分は使っちゃうからな。
その他

最近、センター払い出しユニットの調子が悪いFT3。



オーシャンはやはり苦手・・・

フォーチュンオーブ3初女神リーチ!当時はあの変なヘルメットかぶってなくて、結構可愛い。ダイレクトチャレンジくれたけど、ダイレクトモート突入→1球目終了で600枚減りました。なんだったんだ、あいつ・・・

ファンタジックフィーバーでかぶり始めたっぽい。



珍しく初代トゥインクルドロップで負けてない。

最近やりだした、コナミのメカスロット「ユーロクイーン」。エンドレスフィーバーっていう大チャンスモードに突入すると、懐かしのDDRの曲(my only shining star)が聴ける(ただし、めちゃくちゃ音量小さい)が、行かないときは、3時間回してもいかない。
ただし、マックスベットが5枚なので、そこまで減りは・・・2000枚減った。※ただし翌日戻してくれたけど。
そして、とにもかくにもジャックポットが出ない。5日間でない。
プログレッシブが1000枚から4000枚弱まで増えちゃっているけど、出ない。

エンドレスフィーバー中の一番惜しかったやつ(白が3つ並ぶとジャックポット)。
現在は、少しだけ復興して137000枚。